

Introduction
The development and maturation of B cells is highly dependent on signals provided by the microenvironment of the lymphatic organs. As B cells move from one developmental stage and niche to the next, the integrin family of adhesion molecules provides important cues for their correct positioning and retention. The integrin adaptor protein Kindlin-3 (encoded by the Fermt3 gene) regulates integrin activity and function in a wide range of hematopoietic cell types. In this study, we aimed to define its precise role in the development and function of the different murine B cell subsets.
Methods
We crossed a Fermt3flox/flox mb1-cre mouse strain (hereforth called K3ΔB mice), harboring a B cell specific Kindlin-3 deletion. B cell subsets in the different lymphoid organs of these K3ΔB mice and control littermates were defined by multicolor flow cytometry. Adoptive transfer, microscopy and real-time flow cytometry were used to analyze the different steps of integrin activation. A co-culture system with OP9 stromal cells and BAFF was used to assess the in vitro differentiation potential of immature progenitors into the different mature B cell subsets. Transcriptional differences between follicular B cells isolated from spleens of K3ΔB- and control mice were assessed by transcriptome array.
Results
In vitro, we found that integrin activation on B cells was induced upon activation of the chemokine receptors CXCR4 and CXCR5 or the B cell receptor. This stimulation triggered adhesion of wild type B cells to integrin ligands under shear flow. The increase of VLA-4 integrin affinity to its ligand substrates during this process could also be calculated from real-time flow cytometrical analyses. In contrast, K3ΔB-derived B cells could not reach high affinity states of integrins and thus failed to adhere on the substrates upon stimulation, despite slight upregulation of chemokine receptors CXCR4 and CXCR5. B cell migration towards the respective chemokines also required Kindlin-3, even in an integrin ligand-free setting.
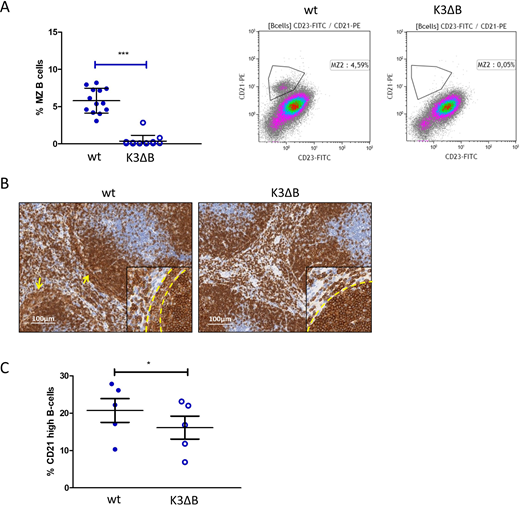
In vivo, Kindlin-3 was required for homing of mature B cells to the bone marrow and to lymph nodes. When further characterizing K3ΔB mice by flow cytometry and immunohistochemistry we observed increased early B cell numbers in the bone marrow. Of note, marginal zone (MZ) B cells in the spleen were completely absent (Figure 1 A+B). We consequently assessed the potential of immature B cells to develop into B cells with high expression of CD21, a marker for MZ B cells, upon their co-culture with OP9 stromal cells in the presence of the B cell survival factor BAFF. While 18% of B cells differentiating from wild type bone marrow displayed high expression of CD21, the percentage of CD21 high cells recovered from Kindlin-3 deficient progenitors was significantly lower (~12%, Figure 1C). Pathways involved in these developmental differences were analyzed by a transcriptome array, revealing increased activity of the B cell receptor pathway in the knockout situation accompanied by higher, NFkappaB and Notch signaling.
Conclusion/Outlook
Whereas our results highlight the importance of Kindlin-3 dependent, integrin mediated cell retention and migration during B cell development they also indicate that Kindlin-3 functions in an integrin-independent manner when regulating cell motility and transcription. The complete lack of MZ B cells in the absence of Kindlin-3 is thus most likely a combination of defective retention in the MZ area and transcriptional alterations favoring the development of transitional B cells into follicular- rather than MZ B cells.
Figure1: B-cell specific Kindlin-3 knockout leads to loss of splenic marginal zone B cells. The percentage of MZ B-cells among total splenic B cells was determined by flow cytometry in K3ΔB mice and wild type (wt) littermates (A). Immunohistochemistry staining of CD19 showed a loss of loosely packed marginal zone B cells (yellow arrows) in the absence of Kindlin-3 (B). B cells were enriched from the bone marrow of K3ΔB mice and wt littermates and cultured on a confluent layer of OP9 cells in the presence of 200 ng/ml BAFF for 72 h. Development of CD21 high/CD23 low B cells was then determined by flow cytometry (C).
Greil:Janssen: Consultancy, Honoraria, Membership on an entity's Board of Directors or advisory committees, Other: Travel, accomodations, expenses, Research Funding; Astra zeneca: Consultancy, Honoraria, Membership on an entity's Board of Directors or advisory committees, Other: Travel, accomodations, expenses, Research Funding; Abbvie: Consultancy, Honoraria, Membership on an entity's Board of Directors or advisory committees, Other: Travel, accomodations, expenses, Research Funding; Daiichi Sankyo, Gilead: Honoraria, Membership on an entity's Board of Directors or advisory committees, Other: Travel, accomodations, expenses, Research Funding; BMS: Consultancy, Honoraria, Membership on an entity's Board of Directors or advisory committees, Other: Travel, accomodations, expenses, Research Funding; F. Hoffmann-La Roche: Consultancy, Honoraria, Membership on an entity's Board of Directors or advisory committees, Other: Travel, accomodations, expenses, Research Funding; BMS/celgene: Consultancy, Honoraria, Membership on an entity's Board of Directors or advisory committees, Other: Travel, accomodations, expenses, Research Funding; Novartis: Consultancy, Honoraria, Membership on an entity's Board of Directors or advisory committees, Other: Travel, accomodations, expenses, Research Funding; MSD Merck: Consultancy, Honoraria, Membership on an entity's Board of Directors or advisory committees, Other: Travel, accomodations, expenses, Research Funding; Takeda: Consultancy, Honoraria, Membership on an entity's Board of Directors or advisory committees, Other: Travel, accomodations, expenses, Research Funding.

This feature is available to Subscribers Only
Sign In or Create an Account Close Modal